過去の研究内容
ゲノムインプリンティングの進化
哺乳類の幾つかの遺伝子は同じ個体内で、父親由来のアレルと母親由来のアレルとで極端に異なる発現量を示す。 このような現象をゲノムインプリンティングという。 メチレーションパターンの違いによって、遺伝子の由来が識別されているらしい。 ゲノムインプリンティングは哺乳類の胎児の成長率を増加させたり減少させたりする遺伝子によく見られる。 例えばIgf2という遺伝子をマウス胎児の体内で強制的に多量に発現させると胚サイズが1.5倍ほどに増加するが、 この遺伝子は正常胚では父親由来の遺伝子からのみ転写される。
Haigは、母親由来の遺伝子と父親由来の遺伝子と間に成長の度合いに関してConfilctが存在することを示し、 それによってゲノムインプリンティングの進化は理解できるとする「綱引き仮説」を提唱した ( Haig, D. & Westoby, M. (1989) Parent-specific gene expression and the triploid endosperm. Am. Nat.134: 147-155)。
胎児は母親の栄養供給によって成長するが、たくさん栄養をもらって大きくなれば良いというものではない。 あまりに栄養を要求しすぎる個体は、兄弟の数を減らしてしまう可能性があり、「適応的」でない。
実は、どれだけ兄弟へ遠慮したらよいかという「最適な」量が、父親由来の遺伝子と母親由来の遺伝子とでは異なる。
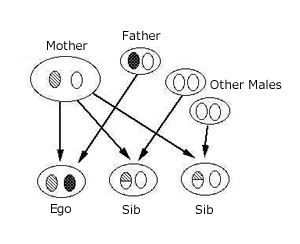
図は遺伝子間の関係を示したもの。 Egoからみて、母親由来の遺伝子がSibにも共有されている確率は0.5だが、父親由来の遺伝子がSibに共有されている確率は、 父親が複数いるときには0.5よりも必ず小さい。父親が全て異なる場合は確率0である。 つまり母親由来の遺伝子の方が、父親由来の遺伝子に比べて、「遠慮した方が良い」のである。
上の議論は直感的に理解しやすいが、遺伝子に主体があるかのような議論であり、 遺伝学に基づいた進化の過程を明らかにしていない。 我々は多変量量的遺伝モデルを用いて、綱引き仮説の妥当性を検証した。 (Mochizuki, A., Takeda, Y. and Iwasa, Y. (1996). The evolution of Genomic Imprinting. Genetics 144: 1283-1295.)
発現量に応じて母親からの栄養供給を増加させるような遺伝子、Embryonic Growth Factorの発現量の進化を考える。 遺伝子は親の性に依存して、発現量を決められると仮定する。 父親由来のときの発現量をx、母親由来のときの発現量をyとして、1つの遺伝子の状態をベクトル(x, y)で表す。
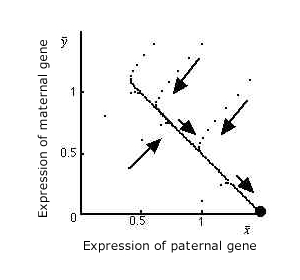
子供はいっせいに産まれて母親の持つ総資源を分け合うと考える”Resource division model”と、 子供は1匹づつ産まれるとする”Sequential care model”の両方のモデルで解析を行ったが、 両者からは定性的に同じ結果が得られた。 すなわち、メスが複数のオスと交尾する時は、 オス由来のアレルからの発現量がメス由来のアレルからの発現量に比べて極端に大きい値に進化する。 結果として、メスが複数のオスと交尾する可能性(Polygamy Rate)がほんの少しでも有れば、 ゲノムインプリンティングは必ず進化することが分かった。
図は成長促進遺伝子の場合。yはほとんど0で、xの方が極端に大きい値に進化する。 成長抑制遺伝子は逆向きに(yの方がxよりも極端に大きく)進化する。