過去の研究内容
シアノバクテリア概日リズムの分子機構
シアノバクテリア時計分子の相互作用を予測する
シアノバクテリアは、概日リズムを示す唯一の原核生物であり、24時間振動の本体として3つの時計遺伝子、KaiA、KaiB、KaiCが同定されている。最近、これらのタンパク質だけを培養したin vitro条件下でも(つまり転写や分解による増減がない環境下でも) KaiCがリン酸化・脱リン酸化の振動を繰り返すことがわかってきた。
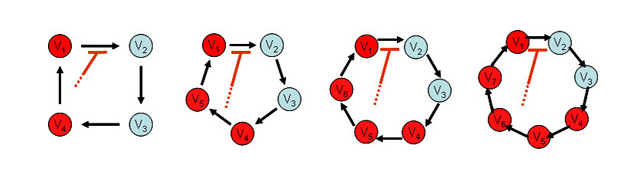
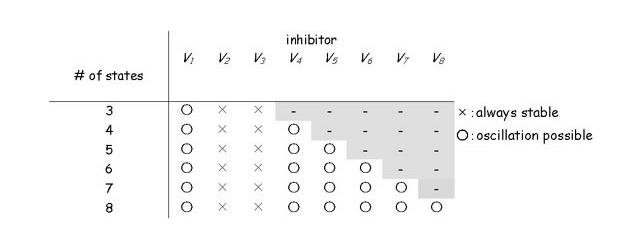
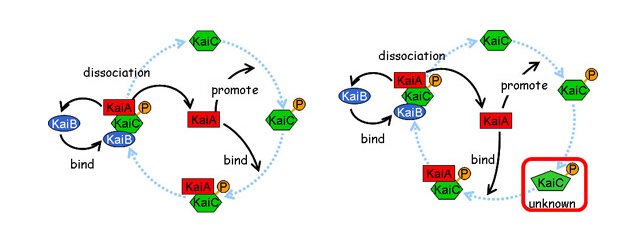
我々は数理モデルを用いて、転写を介さないKaiCのリン酸化振動の機構を明らかにした。タンパク質の状態遷移については、その概要が実験的に明らかにされている。まず、これらをもっとも簡単な形で取り込んだ4変数モデルを解析し、既知の状態遷移だけでは絶対に振動が起こらないことを示した。そこで数理モデルを一般化し、そもそも分子が保存される系において、状態分布の振動が起きるための条件を、数学的に求めた。その結果、状態遷移速度にフィードバックが働く必要性に加えて、フィードバックを受ける反応と与える因子が、一定の状態数以上離れていることが必要だとわかった。ここで得た結果を基に、既知の状態遷移を再検討したところ、タンパク質状態遷移過程に関して、新たな予測が得られた。リン酸化振動が起きるためには、「KaiCがリン酸化された後、KaiAとの複合体を形成する前に、未知の状態を経ていること」が必要である。この予測のもとに「複数リン酸化状態モデル」を構築し、KaiCのリン酸化振動を再現した(Takigawa-Imamura, H., Mochizuki, A. (2006) Predicting regulation of the phosphorylation cycle of KaiC clock protein using mathematical analysis. J Biol. Rhythms 21, 405-416.)。最近、この予測にまさに対応するようなリン酸化状態遷移が、大阪大学、名古屋大学の研究グループにより報告された。
シアノバクテリア概日リズムの分子機構
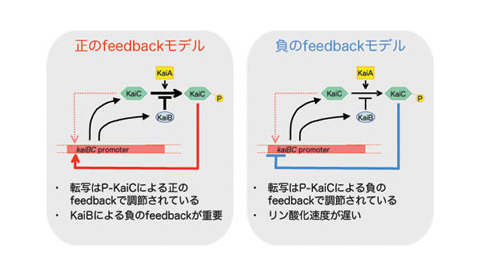
シアノバクテリアは、時計遺伝子の発現振動による概日リズムをもつ細菌であることが知られています。この分子振動は、これまでの概日リズムとは、異なるメカニズムによって実現されている可能性があり、現在実験的研究が熱心に進められています。
我々は数理モデルを駆使することで、実験に先駆けて制御メカニズムの予測を打ちたてようと、取り組んでいます。それは、以下に述べる方法論によって、可能になります。まず、既知の情報を大枠として、未知の機構についてありとあらゆる可能性を取り込んだ数理モデルを、構築します。これを詳細に解析し、振動が起きる条件を数理的に決定します。 概日リズム振動が起きるための必要条件が理論的に定まれば、それは現実の生物でも同様に働いているはずであり、実際のメカニズムの予測に繋がるでしょう (Takigawa-Imamura, H., Mochizuki, A. (2006) Transcriptional Autoregulation by Phosphorylated and Non-Phosphorylated KaiC in Cyanobacterial Circadian Rhythms. J. theor. Biol. 241, 178-192. )。